Il existe une diversité très importante de mécanismes de résistance aux PPP. Ces mécanismes peuvent agir aux différentes étapes de l’action d’un PPP : pénétration dans le bio-agresseur, transport jusqu’au site d’action, accumulation au site d’action, interaction avec le site d’action (en général par fixation sur celui-ci), et effets collatéraux de l’interaction avec le site d’action (par exemple, génération de composés toxiques pour la cellule). Les mécanismes de résistance peuvent être classés en deux catégories : les résistances liées à la cible (RLC) et les résistances non-liées à la cible (RLNC). Les premières aboutissent à une réduction de l’action du PPP sur sa cible physiologique. Les secondes causent une réduction des quantités de PPP atteignant leur cible ou réduisent les effets néfastes de l’action du PPP.
La connaissance des mécanismes de résistance n’est pas seulement importante d’un point de vue théorique ; elle est également un élément essentiel dans le choix d’une stratégie de lutte contre les phénomènes de résistance (voir « Stratégie de gestion des résistances »). Nous présenterons ici les principaux types de mécanismes de résistance en suivant l’ordre des étapes de l’action du PPP.
Ce type de mécanisme permet à un bio-agresseur de survivre aux traitements en évitant d’être en contact avec le PPP (figure 7a, 1). Cet évitement est la conséquence de la sélection d’individus présentant des particularités comportementale (insectes, rongeurs) : les individus résistants vont, par exemple, évoluer vers une répulsion vis-à-vis du PPP. Ils auront donc tendance à éviter le contact avec celui-ci. Une autre possibilité est l’évolution de particularités phénologiques (champignons, insectes, adventices). La conséquence sera une désynchronisation vis-à-vis des traitements : le stade sensible n’est plus ou est peu exposé aux PPP. Pour les adventices, il existe des cas de levée retardée des semences, ce qui permet l’émergence des plantules après l’application des traitements de pré-semis (résistance par esquive). Dans le cas des insectes, certains stades larvaires peuvent être moins sensibles à un PPP que d’autre. Une modification de la vitesse de développement peut donc aboutir à la présence des stades les moins sensibles lors du traitement. Ce type de résistance est principalement décrit chez les insectes et les adventices. On peut supposer qu’il existe également chez les champignons, même si cela n’a pas été étudié de manière formelle.
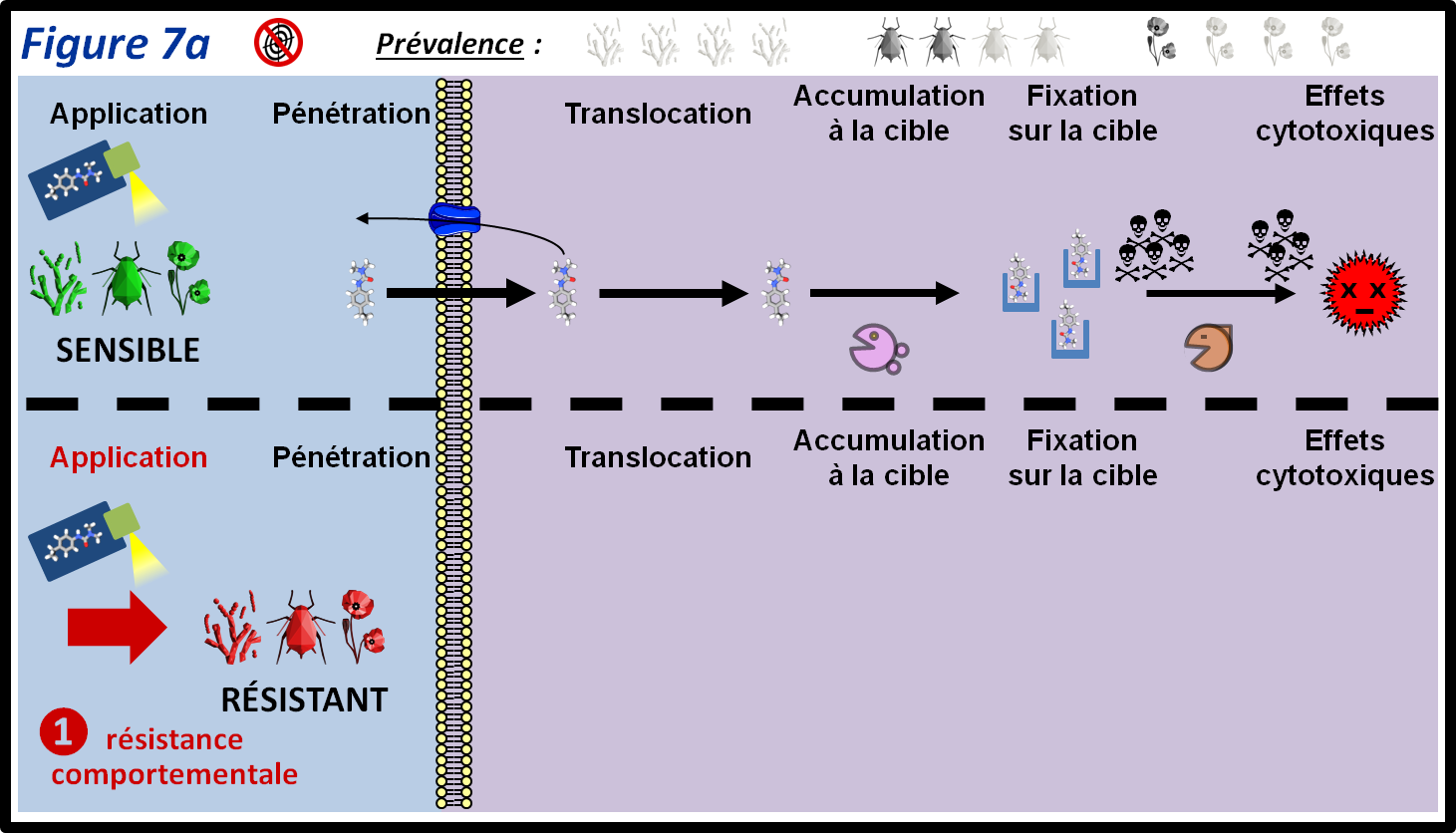
figure 7a : La résistance comportementale ou phénologique permet aux bio-agresseurs d’éviter d’être exposé au PPP. Dans le cas des insectes, il peut s’agir d’effet répulsif de la molécule utilisée. Pour les adventices, une levée tardive peut permettre d’éviter l’exposition au traitement. Les symboles qui représentent les trois grandes classes de bio-agresseurs permettent d’indiquer si le mécanisme de résistance est plus ou moins fréquent au sein de ces classes. La moitié supérieure de la figure montre le parcours du PPP au sein des individus sensibles. Les différentes épaisseurs des flèches représentent les différentes quantités de PPP qui passent d’une étape à l’autre.
Ce type de mécanismes réduit la pénétration des PPP dans l’organisme des bio-agresseurs (figure 7b, 2). Ce sont souvent des modifications de l’épiderme qui aboutissent à une réduction de la pénétration des molécules dans les cellules. Ces évolutions consistent par exemple en une pilosité plus importante empêchant le contact des gouttelettes de PPP avec l’épiderme, une cuticule plus épaisse ou un changement de la composition de celle-ci la rendant plus « imperméable » au PPP.
Ces modifications sont parfois limitées à certaines parties du bio-agresseur. On peut par exemple citer l’évolution de l’épaisseur de la cuticule des pattes des moustiques qui leur confère une résistance aux moustiquaires imbibées d’insecticides. Le port de la plante chez les adventices peut quant à lui modifier la surface exposée lors de l’épandage et/ou réduire l’adhérence des gouttelettes de PPP, réduisant du même coup la quantité de PPP pénétrant dans l’adventice.
Ce type de résistance serait assez commun chez les adventices et les insectes, mais confère des facteurs de résistance faibles. Il est donc souvent associé à un ou plusieurs autre(s) mécanisme(s) chez les individus résistants. La modification de la composition ou de l’épaisseur de la paroi des champignons est plus rare.

figure 7b : La réduction de la pénétration est un mécanisme de résistance décrit chez les trois grandes catégories de bio-agresseurs. La quantité de matière active qui pénètre dans l’organisme est réduite, par exemple par une épaisseur plus importante de la cuticule chez les insectes ou les adventices. Les symboles qui représentent les trois grandes classes de bio-agresseurs permettent d’indiquer si le mécanisme de résistance est plus ou moins fréquent au sein de ces classes. La moitié supérieure de la figure montre le parcours du PPP au sein des individus sensibles. Les différentes épaisseurs des flèches représentent les différentes quantités de PPP qui passent d’une étape à l’autre.
L’efflux (figure 7c, 3) est un type de résistance assez spécifique des champignons. Il consiste, à l’aide de pompes transmembranaires, à excréter les molécules toxiques pour le bio-agresseur, dont les PPP, à l’extérieur du cytoplasme des cellules. Ce type de résistance est probablement répandu chez les champignons phytopathogènes. Un exemple est le phénotype « MDR » (pour Multi Drug Resistance ou résistance multidrogues ; voir plus haut) chez Botrytis cinerea. Comme son nom l’indique, la MDR confère une résistance à de multiples fongicides. La MDR est liée à une mutation (résistance monogénique) induisant la surexpression d’un transporteur membranaire.
Chez les adventices, il existe des cas de résistance par translocation modifiée du PPP. Un exemple notable est un mécanisme de résistance au glyphosate : au lieu d’être transporté vers la localisation de sa cible (les tissus en croissance), celui-ci est convoyé vers l’extrémité des feuilles et les hydatodes (tissus sécréteurs). Ainsi, les dégâts causés par le glyphosate seront circonscrits à une petite partie de l’adventice qui survivra au traitement. De façon similaire, une réduction du transport d’herbicides auxiniques comme le 2,4-D vers leur cible est en cause dans la résistance d’adventices à ces herbicides.
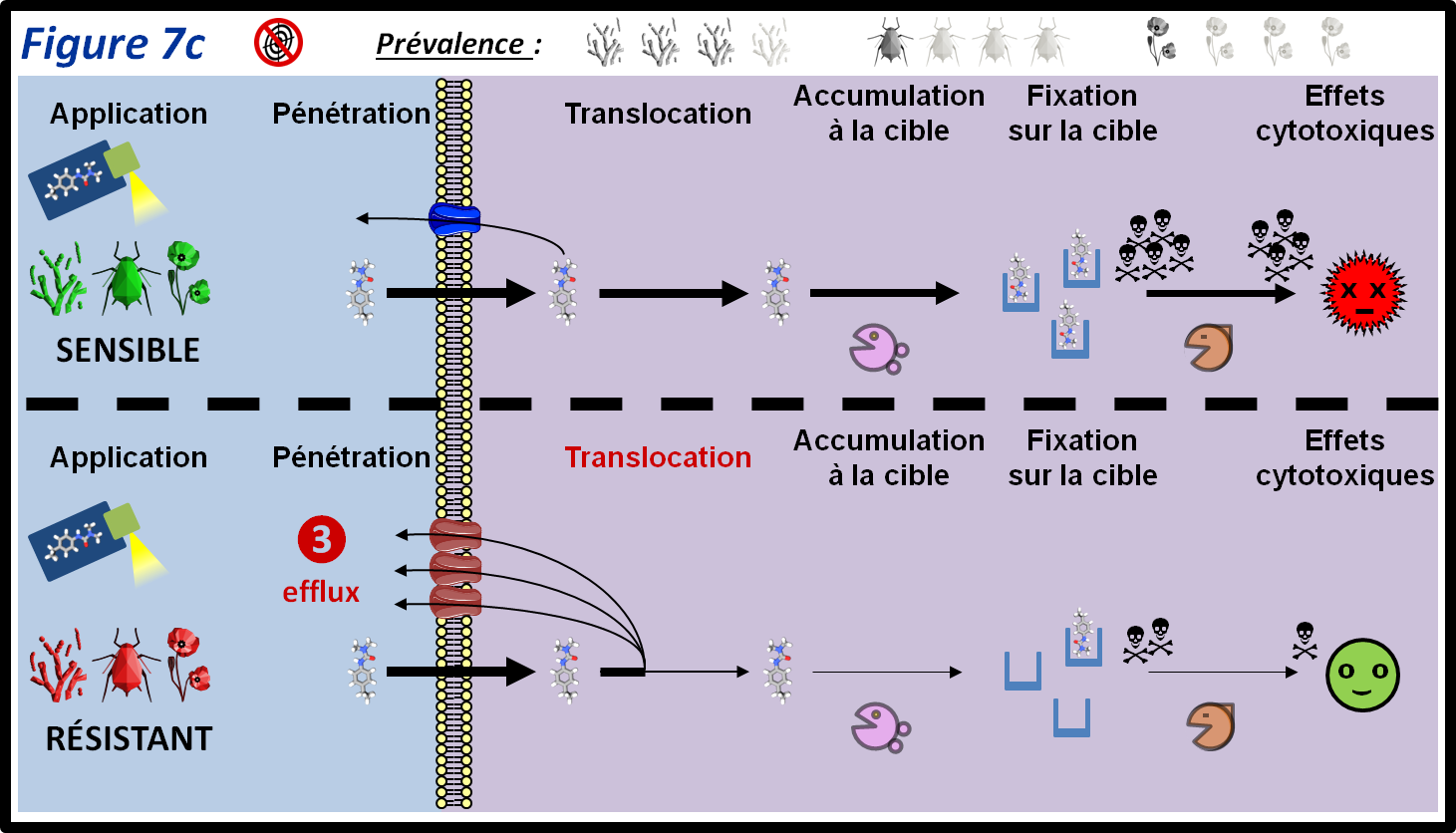
figure 7c : Des systèmes d’efflux ou des modifications du transport des substances ayant pénétré dans l’organisme du bio-agresseur permettent de réduire leur concentration dans l’organisme, ce qui a pour conséquence que le PPP n’est plus efficace. Les symboles qui représentent les trois grandes classes de bio-agresseurs permettent d’indiquer si le mécanisme de résistance est plus ou moins fréquent au sein de ces classes. La moitié supérieure de la figure montre le parcours du PPP au sein des individus sensibles. Les différentes épaisseurs des flèches représentent les différentes quantités de PPP qui passent d’une étape à l’autre.
Il existe différents types de phénomènes de séquestration. Cette séquestration peut être intracellulaire (par exemple au sein des vacuoles, ce qui isole le PPP du reste de la cellule et facilite sa dégradation) ou moléculaire (par complexation avec une protéine ou un enzyme) (figure 7d, 4 et 5, respectivement). Ainsi, un mécanisme de résistance au glyphosate ou au paraquat chez certaines adventices consiste en la séquestration de ces PPP dans les vacuoles. Ce type de mécanismes de résistance semble assez peu fréquent chez les adventices et les champignons, mais assez répandu chez les insectes (séquestration moléculaire).
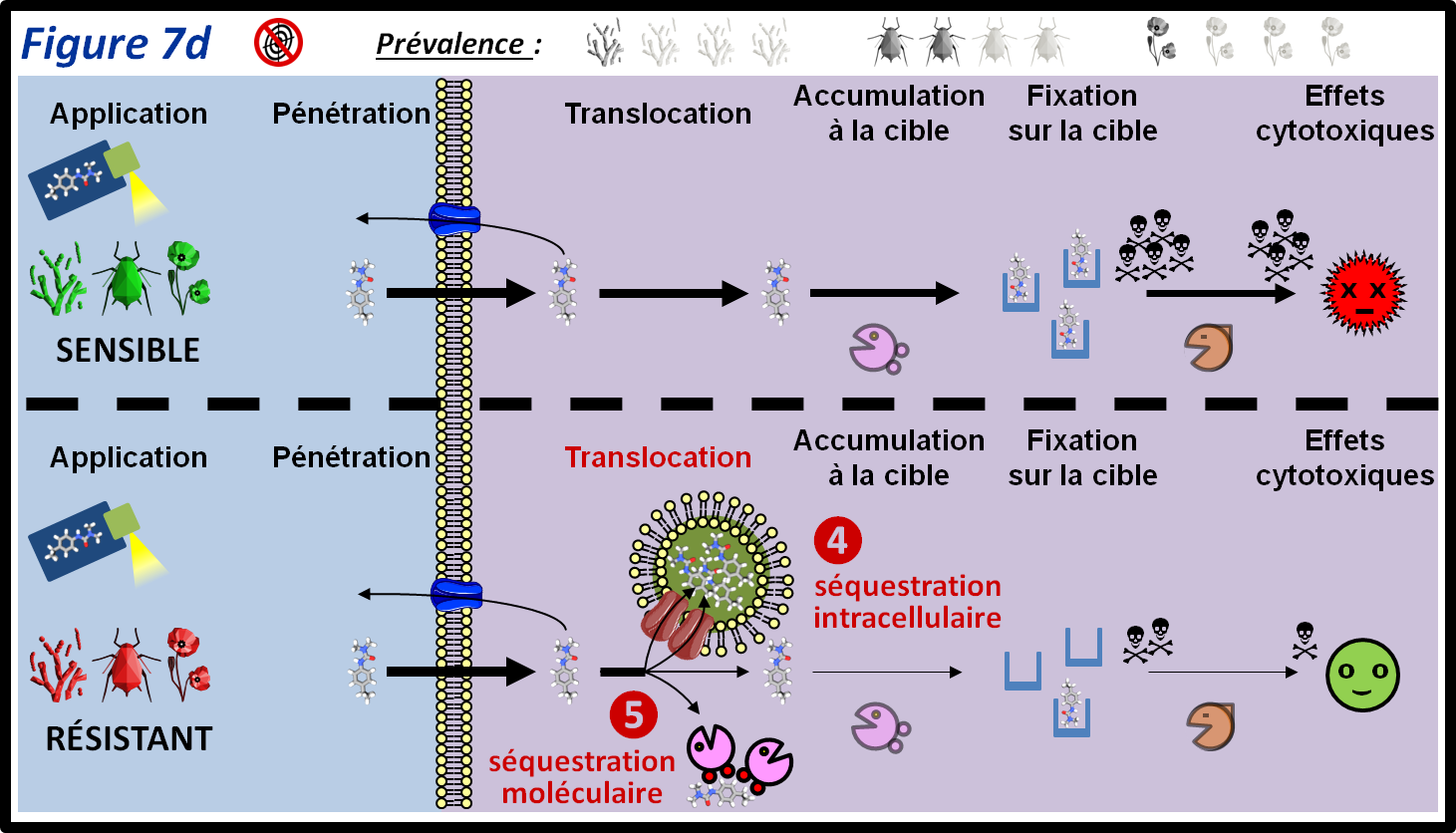
figure 7d : Des mécanismes de séquestration intracellulaire ou moléculaire permettent chez des individus résistants de réduire la concentration du PPP dans les cellules pour que celui-ci n’ai plus un effet létal sur le bio-agresseur. Les symboles qui représentent les trois grandes classes de bio-agresseurs permettent d’indiquer si le mécanisme de résistance est plus ou moins fréquent au sein de ces classes. La moitié supérieure de la figure montre le parcours du PPP au sein des individus sensibles. Les différentes épaisseurs des flèches représentent les différentes quantités de PPP qui passent d’une étape à l’autre.
Toute substance « étrangère » (xénobiotique) pénétrant dans un bio-agresseur y est soumise à une dégradation (métabolisation ou détoxication) plus ou moins rapide due à une ou des voies métaboliques. Une détoxication exacerbée qui confère une résistance à un ou des PPP peut être obtenue par sélection d’individus possédant des enzymes de détoxication plus efficaces et/ou surexprimant de tels enzymes (figure 7e, 6 et 7, respectivement). Ce type de mécanisme est très fréquent chez les insectes et les adventices. Il existe également chez les champignons, mais est peu fréquemment décrit. Un bon exemple de ce type de résistance est la surexpression d’une enzyme (une mono-oxygénase à cytochrome P450) chez le puceron vert du pêcher (Myzus persicae) qui confère une résistance à une famille insecticide : les néonicotinoïdes. Des enzymes de type mono-oxygénase à cytochrome P450, ou glutathion-S-transférases sont en cause dans la dégradation exacerbée de nombreux herbicides chez les adventices.
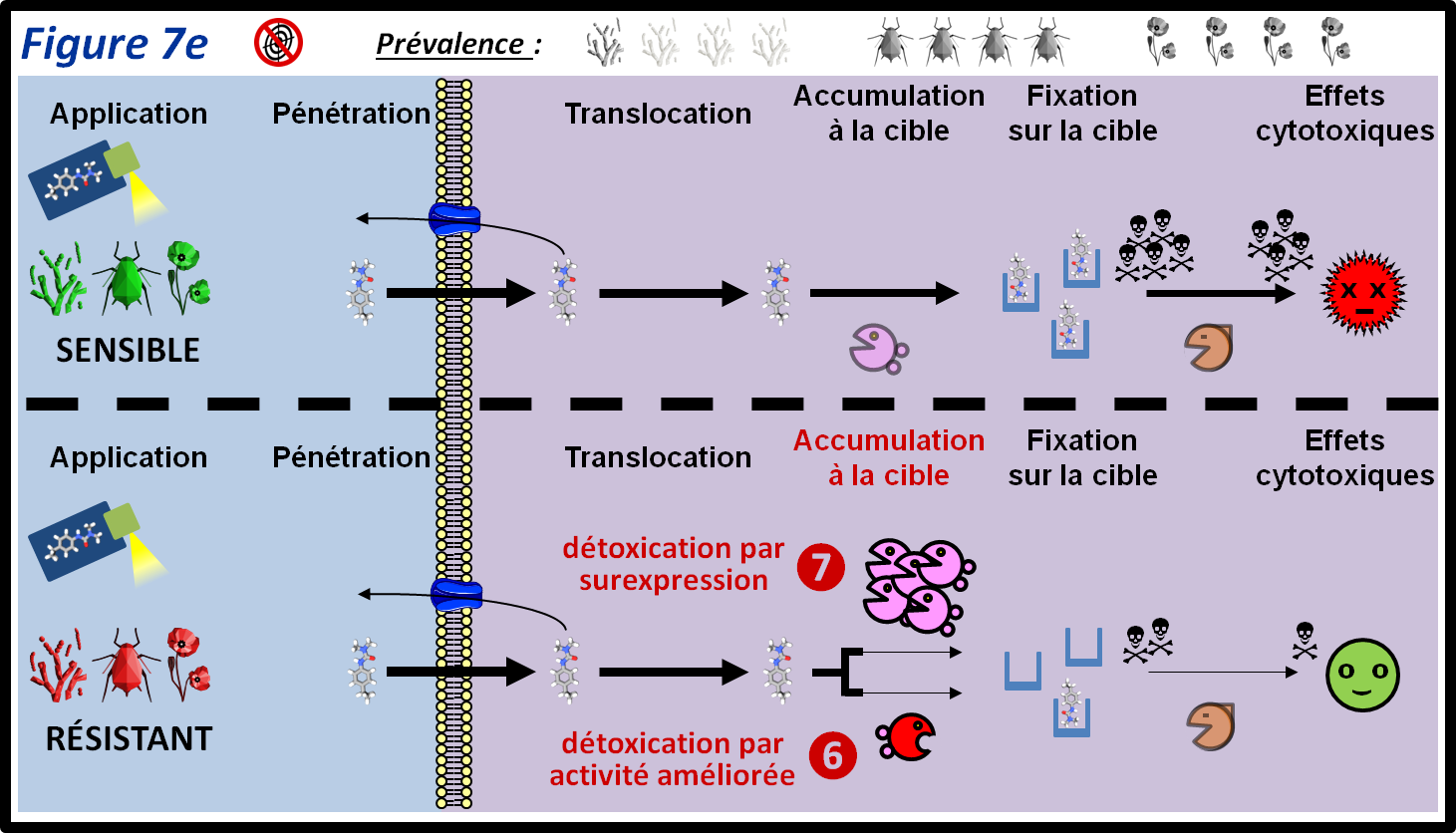
figure 7e : Un mécanisme, très répandu en particulier chez les insectes et les adventices, est la dégradation exacerbée de PPP (détoxication). Celle-ci peut être la conséquence d’une surexpression d’enzymes également présents chez les individus sensibles, et/ou de la présence d’enzyme(s) mutant(s) ayant une activité de dégradation du PPP accrue. La détoxication exacerbée permet de réduire la concentration du PPP à des seuils non létaux. Les symboles qui représentent les trois grandes classes de bio-agresseurs permettent d’indiquer si le mécanisme de résistance est plus ou moins fréquent au sein de ces classes. La moitié supérieure de la figure montre le parcours du PPP au sein des individus sensibles. Les différentes épaisseurs des flèches représentent les différentes quantités de PPP qui passent d’une étape à l’autre.
Ce type de mécanisme de résistance ne modifie pas la sensibilité de la cible à une substance active, mais la quantité de cible présente dans un bio-agresseur. En effet, plus la cible d’une substance active sera présente en grande quantité dans un individu, plus il faudra une dose élevée de substance active pour tuer celui-ci, et donc plus il sera résistant (figure 7f, 8). Ce type de mécanisme revient à « diluer » la substance active dans une plus grande quantité de cible. Il peut être dû à une surexpression du gène de la cible ou à la présence de nombreuses copies du gène codant pour cette cible dans les individus résistants (cas de la résistance au glyphosate chez les adventices). La résistance par surexpression de la cible a été identifiée dans les trois grandes catégories de bio-agresseurs, et en particulier chez les champignons.
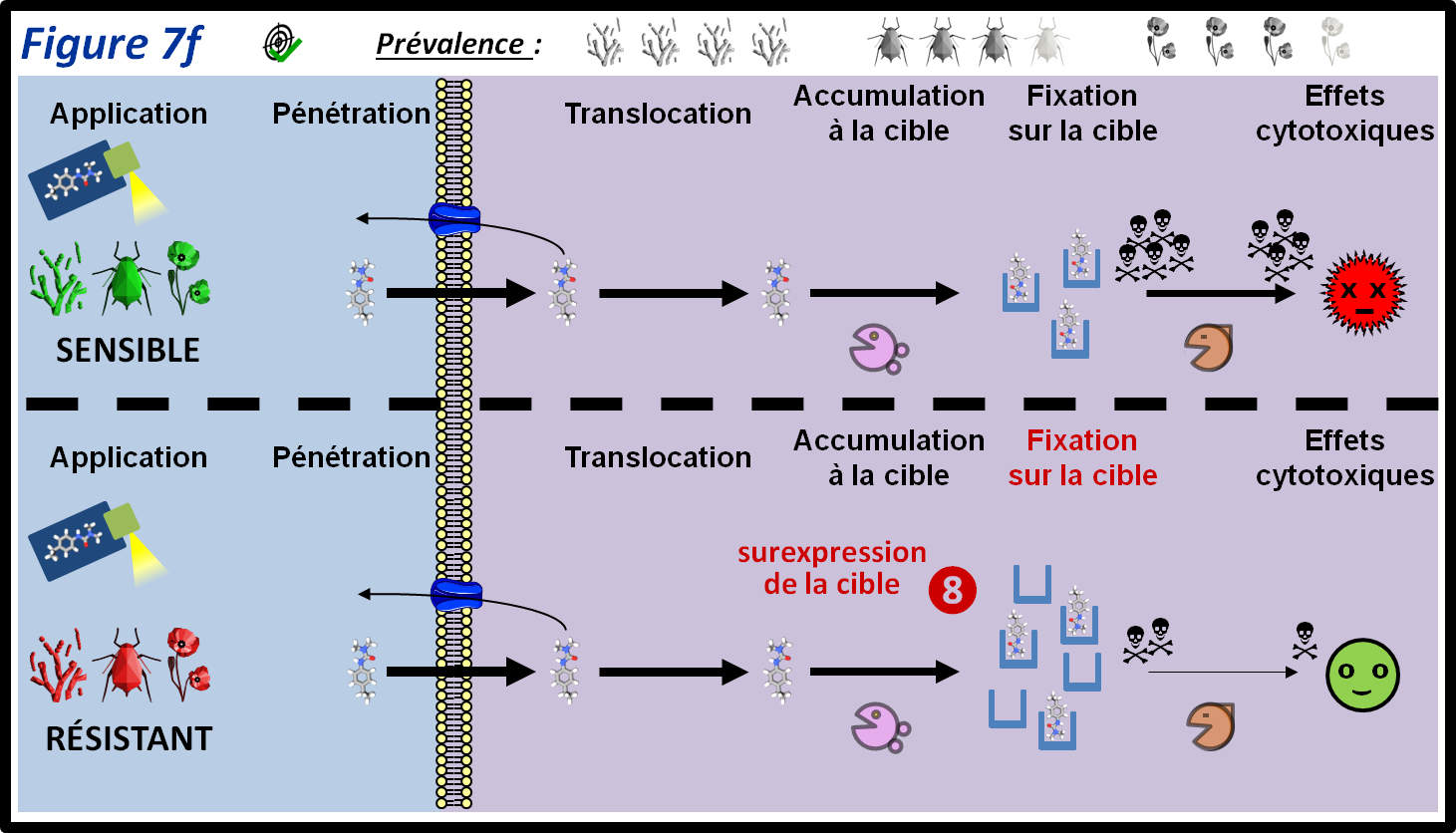
figure 7f : La résistance par surexpression de la cible (RLC) est signalée chez tous les types de bio-agresseurs. La surexpression des protéines ciblées par une substance active permet de « diluer » l’effet de celle-ci, permettant la survie des individus résistants. Les symboles qui représentent les trois grandes classes de bio-agresseurs permettent d’indiquer si le mécanisme de résistance est plus ou moins fréquent au sein de ces classes. La moitié supérieure de la figure montre le parcours du PPP au sein des individus sensibles. Les différentes épaisseurs des flèches représentent les différentes quantités de PPP qui passent d’une étape à l’autre.
Une clef de l’efficacité d’une substance est sa capacité à se fixer efficacement sur sa cible (son affinité pour sa cible). La résistance par mutation de cible est due à une mutation dans le gène codant pour la cible qui cause une modification de la structure tridimensionnelle de celle-ci (figure 7g, 9). La résistance par mutation de cible confère souvent une résistance forte à plusieurs substances agissant sur cette cible (facteur de résistance élevé) mais cela n’est pas systématique : la modification de l’affinité de la substance pour la cible mutée peut-être mineure, ce qui entrainera une faible résistance, voire aucune résistance à certaines substances. Des mutations de cible causant une résistance à certaines substances peuvent même entraîner une hypersensibilité à d’autres substances agissant sur la même cible (herbicides et fongicides inhibiteurs de la polymérisation des microtubules).
La résistance par mutation de cible est le plus fréquemment causée par une seule mutation dans le gène codant pour la cible. Toutefois, il existe des cas d’accumulation de mutations dans des gènes codant pour la cible de fongicides (triazoles) ou d’herbicides (inhibiteurs de l’acétolactate-synthase). En général, le facteur de résistance et le nombre de substances incluses dans le spectre de résistance croisée augmentent avec l’accumulation des mutations.
La résistance par mutation de cible est présente chez les trois grandes catégories de bio-agresseurs. Elle est fréquemment identifiée chez les insectes et les adventices, et très fréquemment chez les champignons.
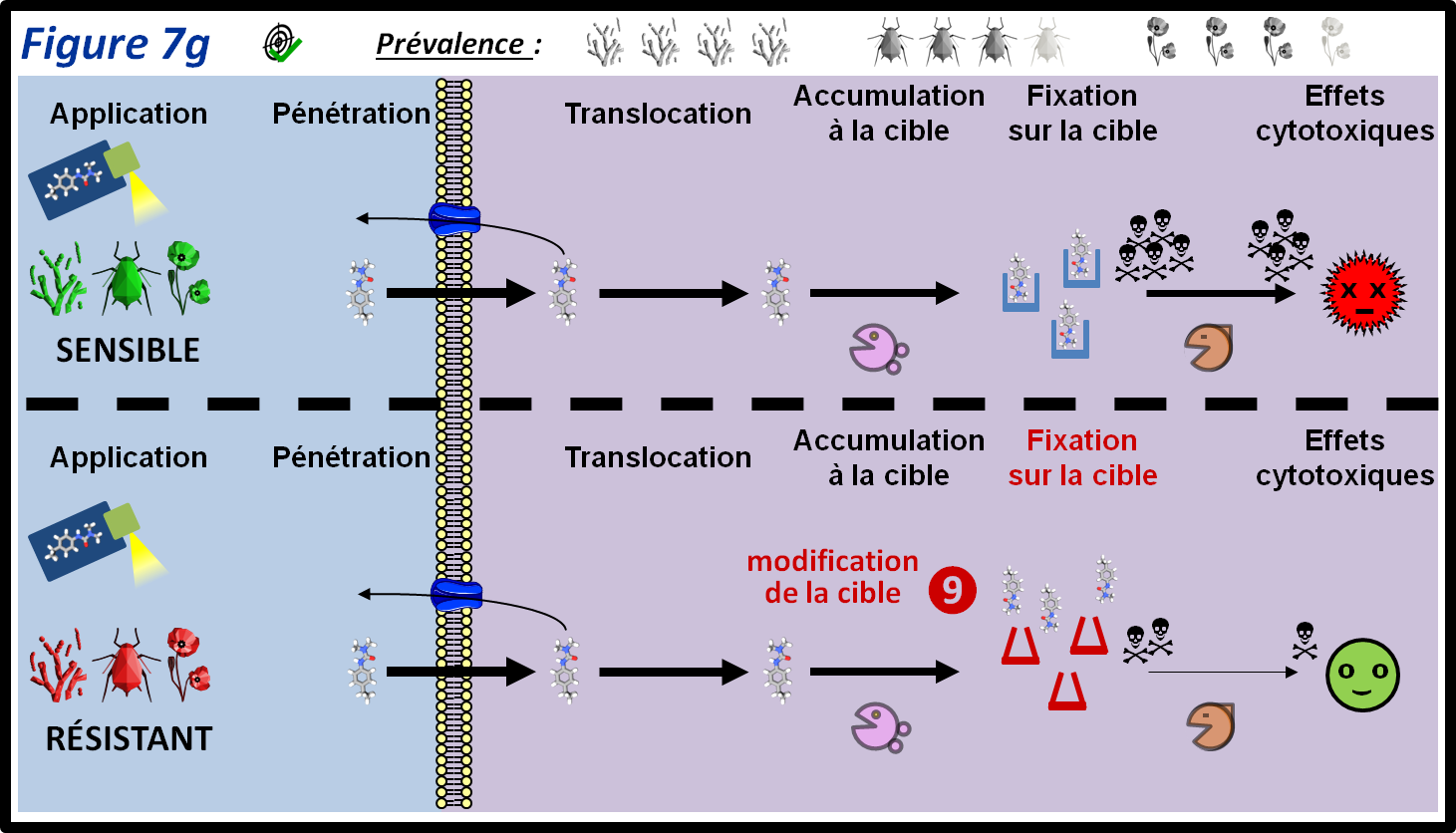
figure 7g : La résistance par mutation de la cible (RLC) est signalée chez tous les types de bio-agresseurs. Une ou des modification(s) structurelle(s) de la protéine cible diminue l’affinité du PPP pour celle-ci, et donc la sensibilité de l’individu. Les symboles qui représentent les trois grandes classes de bio-agresseurs permettent d’indiquer si le mécanisme de résistance est plus ou moins fréquent au sein de ces classes. La moitié supérieure de la figure montre le parcours du PPP au sein des individus sensibles. Les différentes épaisseurs des flèches représentent les différentes quantités de PPP qui passent d’une étape à l’autre.
Chez les individus chez lesquels ce type de mécanisme existe, le PPP exerce l’effet attendu sur sa cible, mais cet effet est compensé par l’activation d’une autre voie métabolique permettant le maintien de la fonction physiologique ciblée par le PPP (figure 7h, 10). Il n’existe à ce jour que quelques exemples de ce type de résistance chez les champignons. Un exemple récent est une résistance du mildiou de la vigne (Plasmopara viticola) vis-à-vis des inhibiteurs du complexe III de la chaine respiratoire des mitochondries (QoI-P, QoI-D et QiI). L’action de ces PPP est compensée par une activité accrue d’un mode de respiration alternatif, l’AOX. Ce type de résistance induit en général une résistance croisée pour toutes les molécules ciblant le même site, la compensation se faisant par une voie biochimique indépendante de la cible originale.
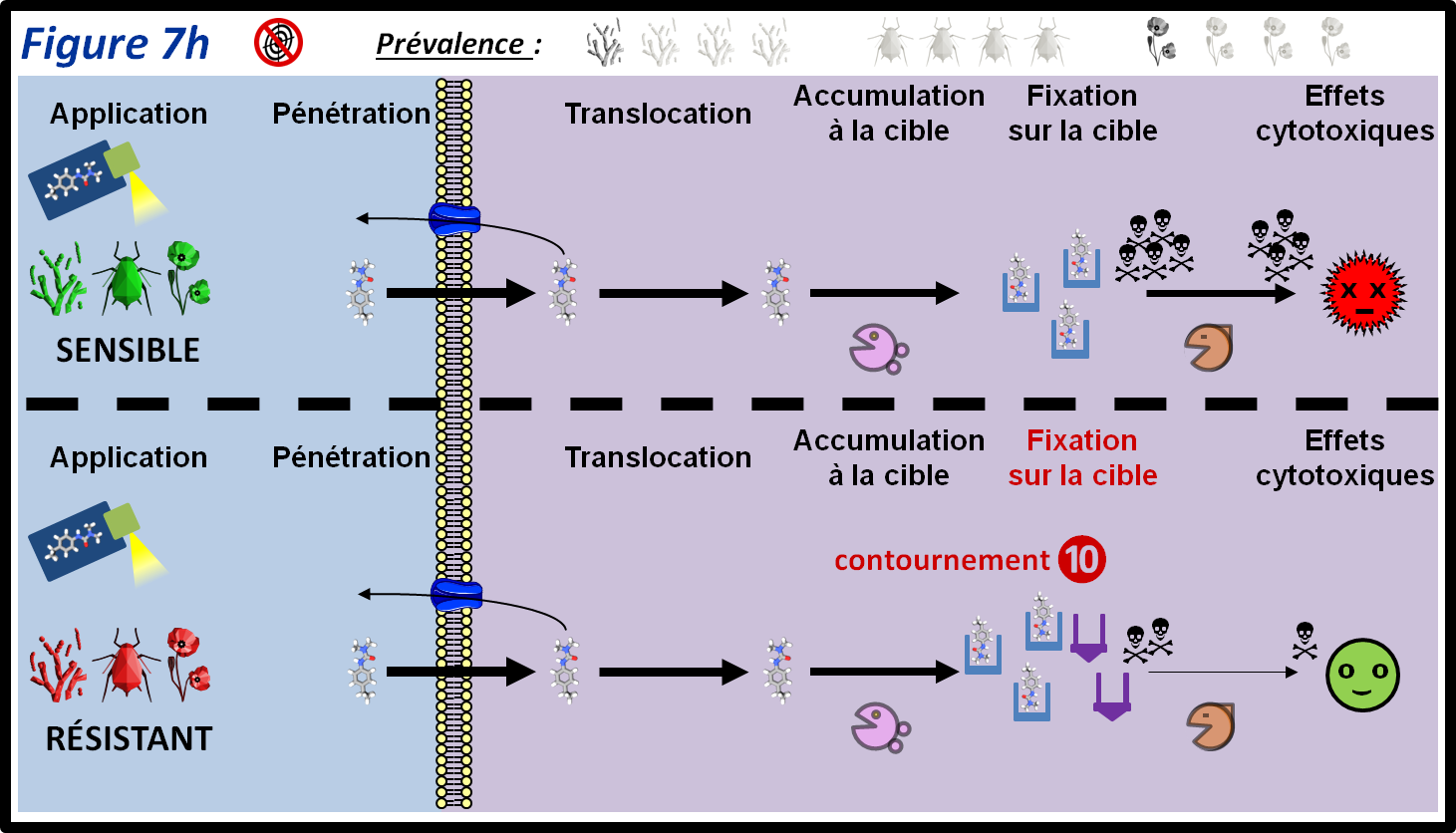
figure 7h : La résistance par contournement est due à l’existence d’une voie alternative qui, chez les individus résistants, compense l’effet du PPP sur sa cible. Les symboles qui représentent les trois grandes classes de bio-agresseurs permettent d’indiquer si le mécanisme de résistance est plus ou moins fréquent au sein de ces classes. La moitié supérieure de la figure montre le parcours du PPP au sein des individus sensibles. Les différentes épaisseurs des flèches représentent les différentes quantités de PPP qui passent d’une étape à l’autre.
L’action d’un PPP sur sa cible peut générer des composés toxiques pour la cellule (peroxydes, radicaux libres…). Les mécanismes de résistance par compensation n’interfèrent pas directement avec le PPP, mais neutralisent ses effets physiologiques secondaires (par exemple, par détoxication des composés cytotoxiques) (figure 7i, 11). Ce type de résistance repose en général sur la surexpression ou la présence de formes plus efficaces d’enzymes de détoxication (par exemple les peroxydases). Ces enzymes peuvent appartenir aux mêmes familles que les enzymes responsables de la détoxication de PPP, mais contrairement à ces derniers, ils n’agissent pas directement sur le PPP. Ce type de résistance a par exemple été démontré chez des adventices contre les herbicides inhibiteurs de l’acétyl-coenzyme A carboxylase.
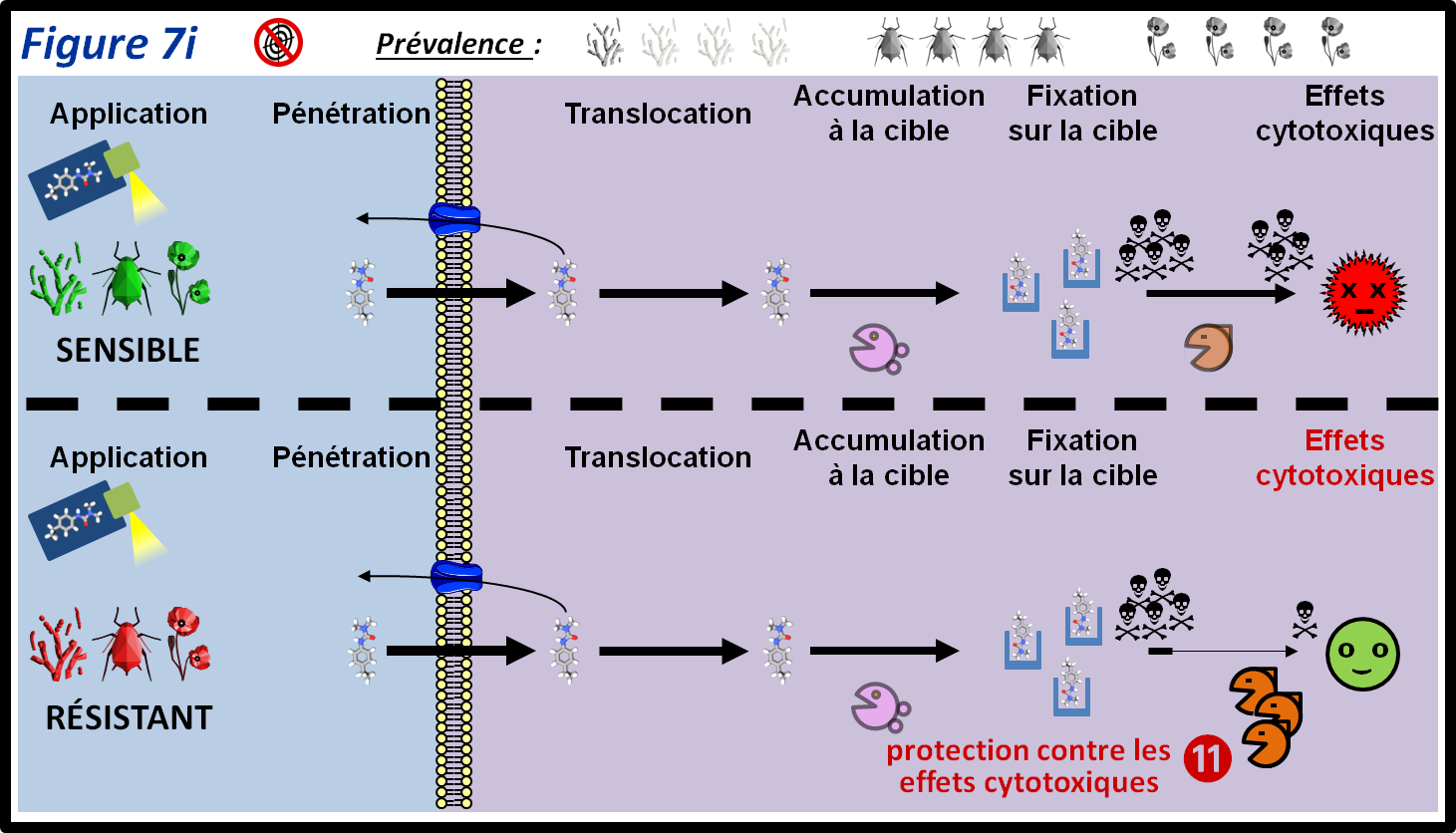
figure 7i : L’action du PPP sur sa cible peut causer la production ou l’accumulation de métabolites cytotoxiques. La résistance par compensation est due à la surexpression et/ou la présence de formes plus efficaces d’enzymes capables de neutraliser ces métabolites. Les symboles qui représentent les trois grandes classes de bio-agresseurs permettent d’indiquer si le mécanisme de résistance est plus ou moins fréquent au sein de ces classes. La moitié supérieure de la figure montre le parcours du PPP au sein des individus sensibles. Les différentes épaisseurs des flèches représentent les différentes quantités de PPP qui passent d’une étape à l’autre.
Synthèse sur les mécanismes de résistances
Les mécanismes qui peuvent conférer une résistance à un ou plusieurs PPP sont extrêmement divers et variés. Ils ne sont bien évidemment pas exclusifs les uns des autres : un individu peut avoir accumulé un ou plusieurs mécanismes lui conférant une résistance à une ou plusieurs substances. La plupart des mécanismes de RNLC sont présents à l’état latent chez les individus sensibles, mais leur efficacité est trop faible pour permettre la survie de ces individus aux doses de PPP appliquées au champ.
Les résistances liées à la cible (ou RLC) peuvent causer des résistances croisées uniquement à des substances actives qui ont la même cible, mais pas forcément systématiquement à toutes. Le facteur de résistance obtenu peut être très élevé, ou non selon les cas. Ceci dépendra à la fois de la substance active et de la mutation. Ce type de résistance étant souvent dû à une mutation sur un seul gène ou sur le promoteur de celui-ci, le spectre de résistance croisé qui lui est associé pourra être intégralement transmis à la descendance.
Les résistances non liées à la cible (RNLC) peuvent concerner des substances ayant des cibles différentes, ce qui les rend extrêmement problématiques du point de vue agronomique. Le facteur de résistance conféré par ce type de résistance est variable. Mais, contrairement à une croyance largement répandue, le facteur de résistance conféré par une RNLC peut être très élevé, en particulier chez les insectes et les adventices. Ainsi, des graminées sont capables de survivre à plus de 14 fois la dose maximale recommandée d’un herbicide inhibiteurs de l’acétyl-coenzyme A carboxylase uniquement grâce à des mécanismes de RNLC. Les RNLC ont une base génétique qui va de la simple mutation à une accumulation de mutations sur différents gènes pouvant modifier à la fois l’expression et l’activité de différentes protéines. C’est pourquoi l’héritabilité de ce type de résistance sera variable, et souvent complexe. Un seul enzyme de détoxication particulièrement efficace pourra être transmis à la descendance tout aussi facilement qu’une résistance liée à la cible. Un mécanisme plus complexe comme la modification d’une voie métabolique pourra reposer sur la nécessiter d’accumuler des mutations sur plusieurs gènes, et aura par conséquence tendance à se dissocier et à ségréger dans la descendance des individus résistants.
Enfin, RLC et RNLC ayant des bases génétiques indépendantes, les deux types de résistance peuvent s’accumuler et coexister dans un même individu. Ainsi, chez les graminées adventices, la RLC aux herbicides inhibiteurs de l’acétolacate-synthase ou de l’acétyl-coenzyme A carboxylase coexiste généralement avec des mécanismes de RNLC dans les individus résistants. De la même manière, chez le carpocapse des pommes des enzymes de détoxifications appartenant à des familles différentes ainsi qu’une mutation dans la cible des insecticides pyréthrinoïdes se sont accumulés dans un même individu devenu « super–résistant » à trois familles insecticides distinctes.